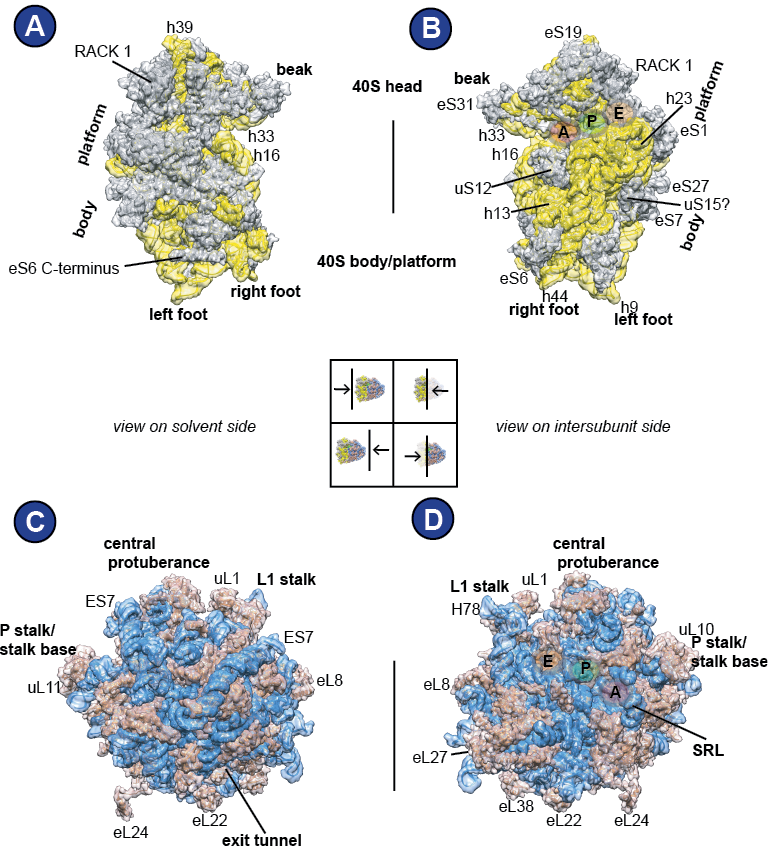
Anatomy of the (mammalian) ribosome
The size of the assembled 80S mammalian ribosome is about 4.3 MDa (Wool, 1979). Both subunits are made of ribosomal RNA (rRNA) and ribosomal proteins (Wool, 1979). Upon joining of the large 60S (50S in bacteria) subunit and the small 40S (30S in bacteria) subunit to the 80S ribosome (70S in bacteria), a functionally important compartment is formed, the so-called intersubunit space (Figure 1), which is one of the main sites of action during protein synthesis. Across it span three tRNA-binding sites, named A (Aminoacyl)-, P (Peptidyl)-, and E (Exit)-sites. Additionally, the ribosome has specific factor binding sites for the interaction with protein factors, like the P-stalk ( Figure 3 C-D).
rRNA is the catalytically active component of the ribosome
The rRNA possesses the catalytic activity to perform peptide bond formation and is the main player in protein synthesis. The large (60S) subunit contains three rRNA molecules: the 28S rRNA, the 5S rRNA and the 5.8S rRNA (Supplemental Figure 1, Supplemental Figure 2). The small (40S) subunit contains only one rRNA molecule, the 18S rRNA (Supplemental Figure 3). The rRNA regions responsible for mRNA-recognition, tRNA-binding and peptidyl-transfer are highly conserved in all kingdoms of life (Gesteland, R.F., Cech, T.R., 1999). Among these conserved regions are the sarcin ricin loop (SRL) on the large subunit, interacting with GTP-hydrolyzing protein factors that catalyze certain steps of translation. In the peptidyltransferase center (PTC), also located on the large subunit, the RNA alone is responsible for catalyzing the formation of the peptide bond (Nissen et al., 2000; Spahn et al., 2000). The small subunit 18S rRNA contains the decoding center (DC), which monitors correct tRNA-anticodon matching to the mRNA codon.
Different from bacterial rRNA, eukaryotic 18S rRNA and 28S rRNA contain several expansion segments, long elements of additional rRNA that are to a great deal responsible for the big difference in size between bacterial and mammalian ribosomes. The function of these expansion segments is not yet clear, and their structural investigation is hindered by their high flexibility and peripheral location, making it very difficult to obtain high-resolution structural information (Ramesh and Woolford, 2016; Yusupova and Yusupov, 2017). There is some evidence, however, that expansion segments may play a role in ribosome biogenesis (Ramesh and Woolford, 2016).
Despite the rRNA’s prominent role in translation, the ribosome would not function without ribosomal proteins. They are important for rRNA folding and assembly and stabilize the tertiary structure of rRNA and the ribosome’s functional centers. There are 33 ribosomal proteins on the 40S and 47 ribosomal proteins on the 60S subunit of the mammalian ribosome. Following a recent convention (Ban et al., 2014) universally conserved proteins are prefixed ‘u’, unique bacterial ones ‘b’, unique eukaryotic ones ‘e’, and unique archaeal ones ‘a’ (Supplemental Figure 4, Supplemental Figure 5).
The three-dimensional shape of the ribosome is optimized for its function
The 40S subunit can be morphologically divided into several regions, named after the 40S subunit’s resemblance in shape to a bird: Looking at it from the solvent site, on top is the 40S ‘head’ with its prominent ‘beak’, below follows the ‘neck’, and the ‘body’ is supplemented by the ‘platform’, ‘shoulder’ and ‘foot’ domains (Figure 3A). In this work, a rough division into two parts will be used: the 40S head (including beak) and the 40S body/platform, comprising the remaining domains. Importantly, the link between the 40S head and 40S body/platform is flexible and allows for intrasubunit motions. Visible from the intersubunit space, there are the three 40S tRNA binding sites that are distributed among the 40S head and 40S body/ platform: A, P, and E ( Figure 3 B). The rRNA residues that constitute the tRNA binding sites are well-conserved (Supplemental Figure 1, Supplemental Figure 2, Supplemental Figure 3).
The large (60S) subunit is characterized by several landmarks; The central protuberance, the P-stalk/stalk base and the L1 stalk ( Figure 3 C-D). From the solvent side, one can see the ribosomal exit tunnel, from which the newly synthesized protein emerges. The solvent side is to a large degree covered by expansion segment ES7, the largest expansion segment of the 28S rRNA ( Figure 3 C). Looking on the 60S from the intersubunit space reveals the A-, P-, and E-tRNA binding sites and the sarcin-ricin loop (SRL) ( Figure 3 D).